Imunobiológia
Obsah
Bunky imunitného systému
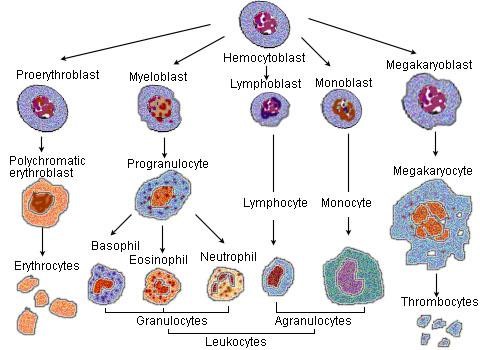
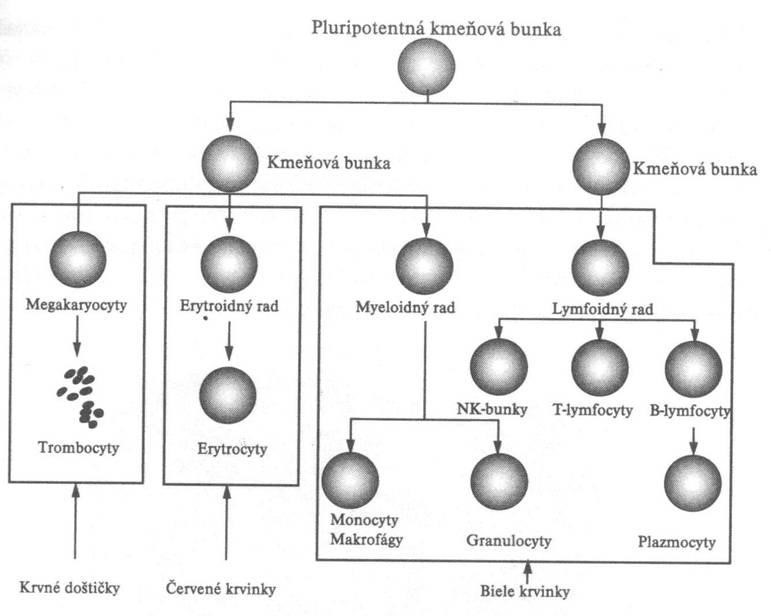
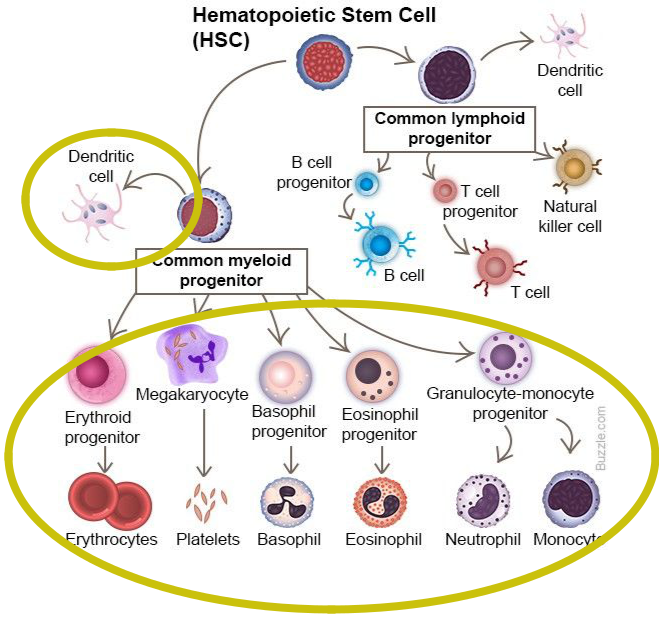
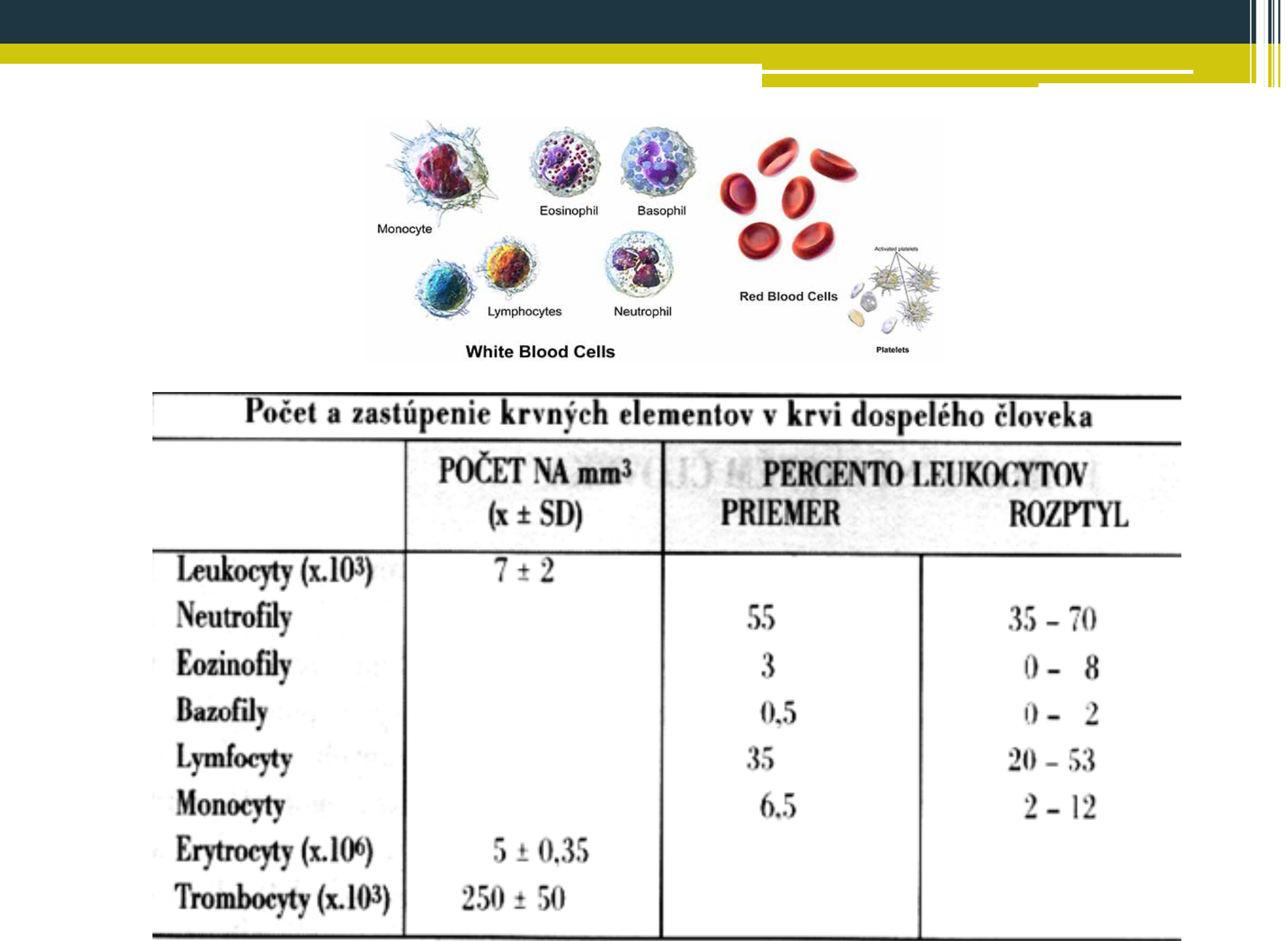
Bunky zúčastňujúce sa imunitných mechanizmov môžeme rozdeliť na bunky patriace bezprostredne do imunitného systému a na pomocné (podporné) bunky. Základnými bunkami imunitného systému sú biele krvinky (leukocyty), vznikajúce z pluripotentných kmeňových buniek, prítomných v kostnej dreni. Tieto bunky sa udržiavajú v kostnej dreni v malom počte po celý život, 1 kmeňová bunka prináleží na 10000 buniek kostnej drene. Pluripotentná kmeňová bunka sa u človeka najprv sa objavuje v žĺtkovom vaku, v 3. týždni migruje do fetálnej pečene, potom do sleziny. Z tohto dôvodu sú pečeň a slezina miestami hematopoézy medzi 3. - 7. gestačným týždňom. Potom sa presúva do kostnej drene, ktorá sa stáva hlavným hematopoetickým orgánom. Do narodenia tvorba krvných buniek v pečeni a slezine prestáva, hematopoéza sa koncentruje do plochých kostí a v dospelosti je sústredená na sternum, bedrové kosti, chrbticu a rebrá. Vo včasnej hematopoéze vzniká z pluripotentnej kmeňovej bunky v dôsledku pôsobenia rastových faktorov myeloidná progenitorová kmeňová bunka a lymfoidná progenitorová kmeňová bunka. Tieto bunky sa diferencujú do prekurzorových buniek, ktoré stratili schopnosť vlastnej sebaobnovy a sú predurčené na vývoj len určitým smerom. Z myeloidného prekurzoru (tzv. myeloidná dráha) vznikajú monocyty, makrofágy, neutrofilné, eozinofilné a bazofilné granulocyty, dendritické bunky, erytrocyty a trombocyty. Z lymfoidného prekurzoru (tzv. lymfoidná dráha) sa diferencujú B lymfocyty, T lymfocyty a NK bunky.
{kind=link}
{kind=link}
Myeloidná dráha
Z myeloidnej dráhy vznikajú monocyty (ktoré cirkulujú v krvi a v tkanivách sa diferencujú na makrofágy) a tri druhy granulocytov: neutrofilné (polymorfonukleárne - PMNL), eozinofilné a bazofilné. Tkanivovou formou bazofilov sú žírne bunky (mastocyty). Do tejto vývojovej vetvy patria aj dendritické (dendritové) bunky. Všetky druhy myeloidných buniek tvoria základ nešpecifickej časti imunitného systému. Väčšina z nich má schopnosť fagocytózy a sú producentmi cytokínov a ďalších rozpustných mediátorov. Z myeloidnej dráhy pochádzajú aj erytrocyty a trombocyty (pomocné alebo podporné bunky), ktoré majú význam pri niektorých imunitných reakciách (najmä pri zápale), ale ich hlavné funkcie sú iné. Dendritové bunky, ale aj monocyty a makrofágy pôsobia ako bunky prezentujúce antigén (APC bunky) pre T lymfocyty a sú aj súčasťou antigénnej špecifickej časti imunitného systému.
{kind=link}
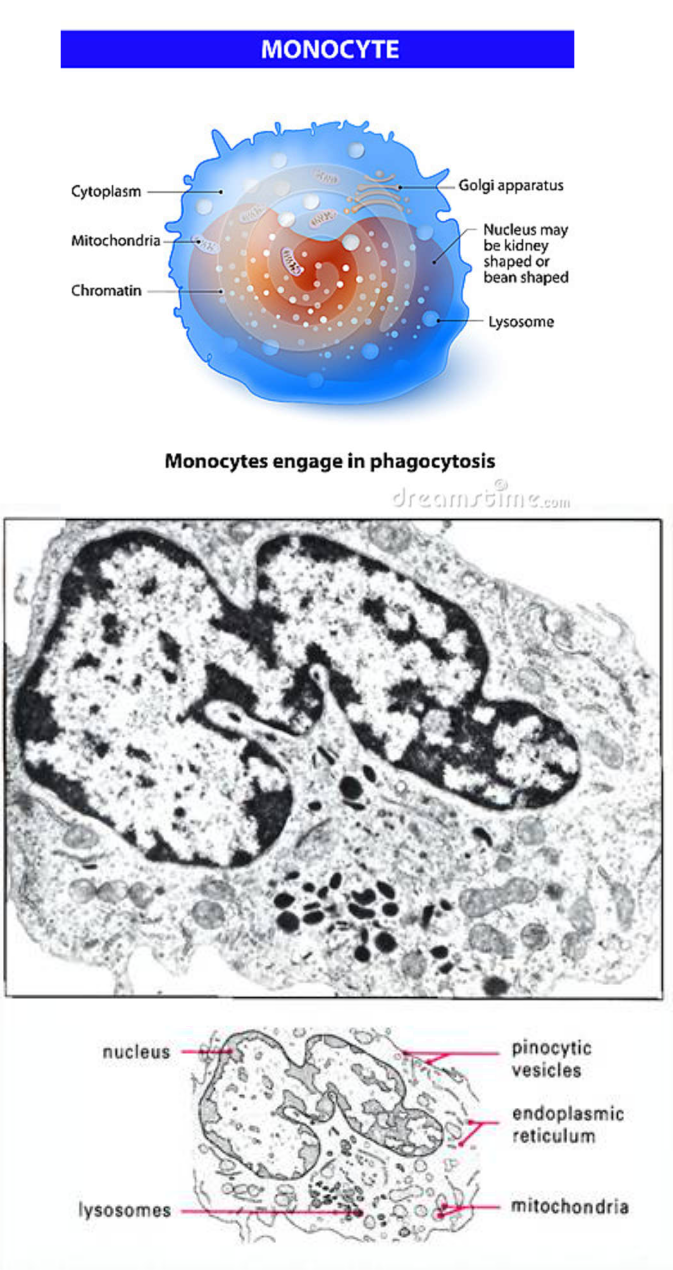
Monocyty
Monocyty sú najväčšími bunkami krvi (priemer 16 - 22 pm), podobné lymfocytom. V periférnej kvi sú zastúpené 2 - 10 % z prítomných leukocytov. Majú oválny alebo okrúhly tvar s veľkým podkovovitým jadrom. V cytoplazme sú početné pinocytové vezikuly, lyzozómové granuly, mitochondrie a granulované endoplazmatické retikulum. Sú súčasťou mononukleárno-fagocytového systému (MPS - Mononuclear Phygocyte system) ako tzv. profesionálne fagocyty. Začiatočná vývojová forma sa nazýva monoblast, mení sa na promonocyt. Promonocyt sa v kostnej dreni delí, počas 6 dní vstupuje do krvi a diferencuje sa na monocyt. Po 3 dňoch monocyty opúšťajú cirkuláciu, usídľujú sa v tkanivách a menia sa na makrofágy (ich životnosť sa predlžuje, premena sa realizuje prestavbou cytoplazmy). Monocyty vtákov sú stavbou podobné vtáčím veľkým lymfocytom s väčším zastúpením cytoplazmy.
{kind=link}
Makrofágy
Makrofágy sú evolučne najstaršími sprostredkovateľmi vrodenej imunitnej reakcie. Aj keď tvoria iba 5 - 10 % populácie leukocytov, zohrávajú centrálnu úlohu nielen pri vrodenej, ale aj špecifickej imunite. Po vzniku prechádzajú niekoľkými štádiami a na rozdiel od neutrofilných granulocytov (neutrofilov) sú pri infekcii schopné deliť sa. Majú tiež vyššiu životnosť a keďže ich odpoveď na infekciu je pomalšia, dominujú v neskorších fázach vrodenej imunitnej reakcie. Majú typické dlhé pseudopódie, ktoré sa podieľajú na tvorbe fagozómu.
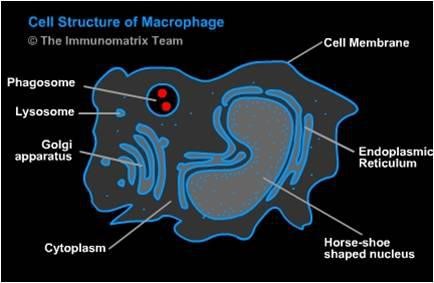{kind=link}
Ďalším rozdielom medzi neutrofilmi a makrofágmi je v schopnosti prezentácie antigénu. Makrofágy a dendritové bunky, na rozdiel od neutrofilov, majú schopnosť na svojom povrchu prezentovať antigény (sú APC bunkami). Fagocytovaný antigén upravia a takto upravený antigénový determinant vystavia na svojom povrchu v spojení s MHC molekulami.
Počas fagocytózy makrofágy syntetizujú a uvoľňujú imunoregulačné látky, napr. cytokíny, zložky komplementu, fragmenty kolagénu, elastín a fibrinogén. Nie sú schopné syntetizovať myeloperoxidázu, tento enzým získavajú z okolia. Z enzýmov syntetizujú hlavne elastázu a kolagenázy, ktoré sa podieľajú na rozklade poškodeného tkaniva, čoho výsledkom je bunkový detritus v zápalových procesoch.
Makrofágy sú schopné sysntetizovať aj rastové faktory pre fibroblasty, čo sa využíva v procesoch hojenia rán, remodelácii tkaniva, v procesoch angiogenézy.
Tkanivové makrofágy sa rozdeľujú na základe tkanivovej, resp. orgánovej príslušnosti na makrofágy: spojivového tkaniva (histiocyty), pečene (Kupfferove bunky), pľúc (alveolárne makrofágy), lymfatických uzlín a sleziny (voľné a usadené makrofágy), kostnej drene (usadené makrofágy), kostí (osteoklasty), seróznych tekutín (pleurálne a peritoneálne makrofágy), kože (histiocyty, Langerhansove bunky) a iných tkanív.
Dendritové bunky
Dendritové bunky (DCs) sú APC bunkami, ktoré zohrávajú dôležitú úlohu pri usmerňovaní imunitnej odpovede. Často sú označované aj ako vlasaté bunky, pre svoje charakteristické pseudopódie. Dendritové bunky sa vyskytujú najmä v slezine a lymfatických uzlinách. Aj keď nemajú schopnosť fagocytózy, sú príbuzné makrofágom. Schopnosť DCs spracovávať a prezentovať širokú paletu antigénov, ktorým je organizmus vystavený, je unikátna. DCs sa okamžite aktivujú pri prieniku patogénu do organizmu. Zároveň však zohrávajú rozhodujúcu úlohu v adaptívnej odpovedi, ktorá sa rozvíja následne. Vzhľadom na to, že aktivácia T lymfocytov závisí od aktivácie dendritových buniek, DCs slúžia ako prvotné rozhranie, spájajúce vrodenú a adaptívnu imunitu. Množstvá dendritových buniek sú denne tvorené v kostnej dreni, vyplavované do cirkulácie a odtiaľ migrujú do tkanív, kde zotrvávajú vo forme nezrelého štádia. Nachádzajú sa takmer vo všetkých tkanivách okrem centrálnej nervovej sústavy.
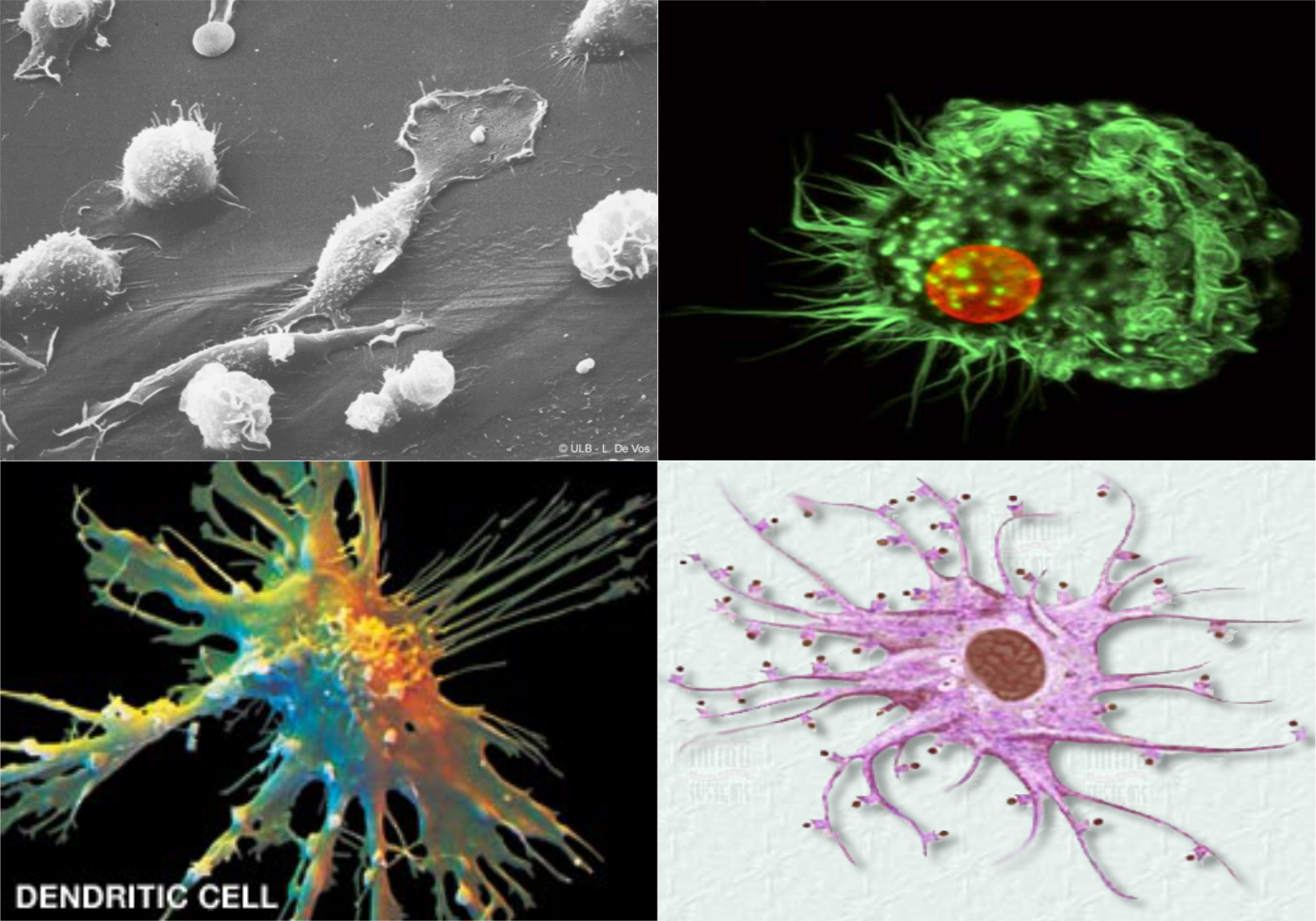{kind=link}
Nezrelé DCs majú vysokú fagocytárnu a endocytárnu kapacitu a neustále monitorujú svoje okolie na zachytenie antigénov potencionálnych patogénov. Po kontakte s antigénmi a expozícii „signálom nebezpečenstva“ (patogény, poškodenie tkaniva, lokálny zápal), DCs dozrievajú a migrujú do lymfatických orgánov. Počas tejto migrácie, DCs podliehajú zreniu, ktoré je charakterizované znížením kapacity vychytávať antigény, zvýšením kapacity spracovania a prezentácie antigénov a zvýšením expresie kostimulačných molekúl pre komunikáciu s lymfocytmi. V sekundárnych lymfatických orgánoch zrelé DCs prezentujú antigény zachytené na periférií naivným a pamäťovým T lymfocytom a indukujú adaptívnu imunitnú odpoveď. Kapacita DCs stimulovať T lymfocyty závisí od stavu zrelosti DCs. Pri absencii silných zápalových stimulov sú pokojové, nezrelé DCs slabými induktormi proliferácie T lymfocytov a pravdepodobne navodzujú toleranciu. Po odovzdaní informácie o patogénoch v sekundárnych lymfatických orgánoch nie je funkcia dendritovej bunky ďalej potrebná pre akútnu odpoveď a DCs pravdepodobne podliehajú apoptóze a zanikajú.
Neutrofilné granulocyty
Neutrofilné granulocyty (polymorfonukleárne leukocyty - PMNL), sú hlavnými zástupcami leukocytov (50 - 70 % u človeka). Bunky majú veľkosť 12 - 15 pm, sú guľatého tvaru a obsahujú charakteristické viaclaločnaté, segmentované jadro (3-5 segmentov) v prípade zrelých buniek a tyčinkovité jadro u nezrelých foriem neutrofilov. V cytoplazme sú početné primárne (azurofilné) granuly, obsahujúce myeloperoxidázu, neutrálne proteázy (elastáza), kyslé hydrolázy (0-glukuronidáza), katepsín B a sekundárne granuly s obsahom lyzozýmu, kolagenázy, laktoferínu, proteínu viažúceho B12. V cytoplazme sú prítomné aj glykogénové zrná ako zdroj glukózy pre anaeróbnu glykolýzu. Tento proces využívajú neutrofily pri zotrvaní v poškodenom tkanive s nízkym obsahom O2, napr. v mieste bakteriálnej infekcie. Významnou vlastnosťou neutrofilov je uplatnenie reaktívnych foriem kyslíka a dusíka v procese likvidácie baktérií, označovanom ako respiračné vzplanutie leukocytov. Neutrofily vznikajú v kostnej dreni, kde sú zadržiavané asi 5 dní, potom sa dostávajú do cirkulácie a na chemotaktické podnety prechádzajú do extravaskulárneho priestoru v miestach zápalových reakcií. Z tohto dôvodu sú primárnymi bunkami zápalového procesu. V tele dospelého človeka ich denne vzniká okolo 1011, pričom majú veľmi krátku životnosť (v priemere okolo 5 hodín). Pokiaľ sa nezúčastňujú fagocytózy a nie sú následne likvidované tkanivovými makrofágmi, dochádza k ich apoptóze. V lymfatických uzlinách sa môžu vytvárať zásoby neutrofilov, ktoré sa v prípade infekcie dajú veľmi rýchlo použiť.
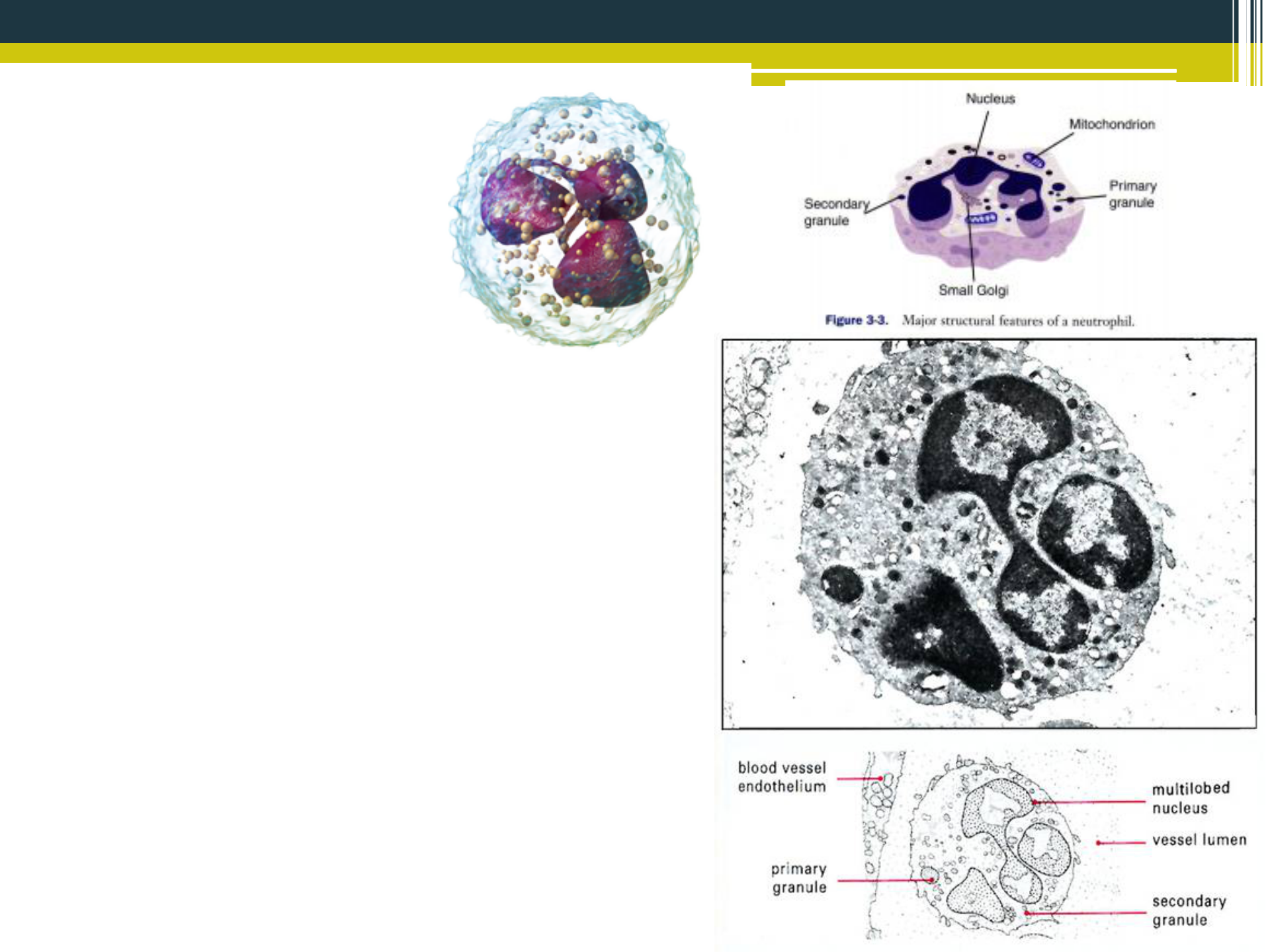{kind=link}
Neutrofilné granulocyty sú základnými bunkami prvej obrannej línie hostiteľa proti patogénnym mikroorganizmom, cudzorodým bunkám a iným látkam. Označujú sa ako profesionálne fagocyty. U vtákov sa neutrofily označujú ako heterofilné bunky. V porovnaní s cicavčími granulami neutrofilov sú vtáčie granuly acidofilné. Vtáčie granuly heterofilov majú vretenovitý tvar (nie okrúhly ako u cicavcov) a jadro je viaclaločnaté.
Eozinofilné granulocyty
Eozinofily veľkosťou a tvarom pripomínajú neutrofily. Majú taktiež viaclaločnaté jadro (najčastejšie 2 segmenty) s početnými špecifickými granulami, obsahujúcimi hlavne kyslú fosfatázu, peroxidázu, histaminázu, hlavný bázický proteín (MBP) a eozinofilový kationický proteín (ECP). Patria medzi profesionálne fagocyty, uplatňujúce sa hlavne v imunitných reakciách proti parazitom (helmintom). Charakteristický prejav endoparazitóz (teanióza, askaridóza, amebiáza) je eozinofília. Parazity sú opsonizované a eozinofily uvoľňujú obsah svojich granúl (MBP a ECP) na povrch opsonizovaných parazitov, čo vyvolá lýzu buniek s následnou smrťou parazita. Prostredníctvom prítomnosti histamináz sa zúčastňujú na tlmení a ukončení zápalových reakcií alergického pôvodu. Vtáčie eozinofily majú eozinofilné granuly a členité jadro, čiže sú zhodné s vtáčími heterofilmi. Eozinofilné granuly sú však okrúhle a sú nevýrazne červené v porovnaní s heterofilnými granulami. Eozinofily dosahujú veľkosť 13-20 pm, v krvnej cirkulácii tvoria 2 - 5 % z prítomných leukocytov.
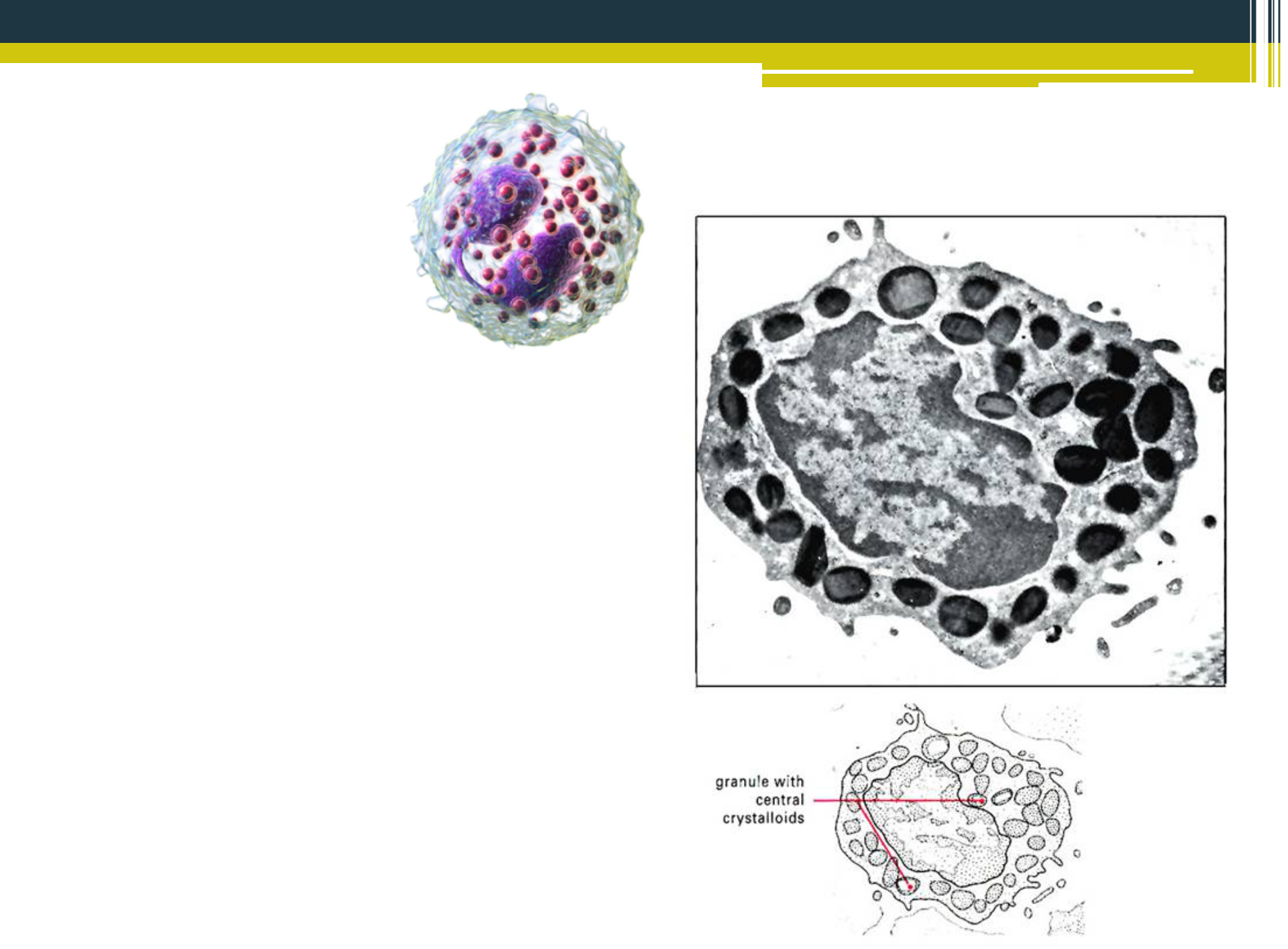{kind=link}
Bazofilné granulocyty
Krvné bazofily sú podobné mastocytom. V cytoplazme sa nachádzajú početné granuly, obsahujúce histamín, bradykinín, serotonín a lyzozómové enzýmy. . Nabunkovej membráne disponujú receptormi (Fc) pre molekuly imunoglobulínu triedy E (Ig E), prostredníctvom čoho sa podieľajú na vzniku alergických reakcií. Po aktivácii receptora s Ig E dochádza k degranulácii bazofilov a vplyvom uvoľnených mediátorov dochádza k lokálnej vaskulárnej a tkanivovej alergickej reakcii. Bazofily podporujú alergické reakcie, pričom eozinofily majú tendenciu tlmiť tieto procesy. Medzi funkciami týchto dvoch typov buniek existuje rovnováha. V krvi sú zastúpené v rozmedzí 0,5 - 2 % zo všetkých leukocytov. Dosahujú veľkosť 13 - 20 ^m. Jadro je veľké pretiahlé, rozdelené na 2 - 3 laloky, prekryté granulami. Bazofily sú nefagocytujúcimi granulocytmi s nepatrným amébovitým pohybom.
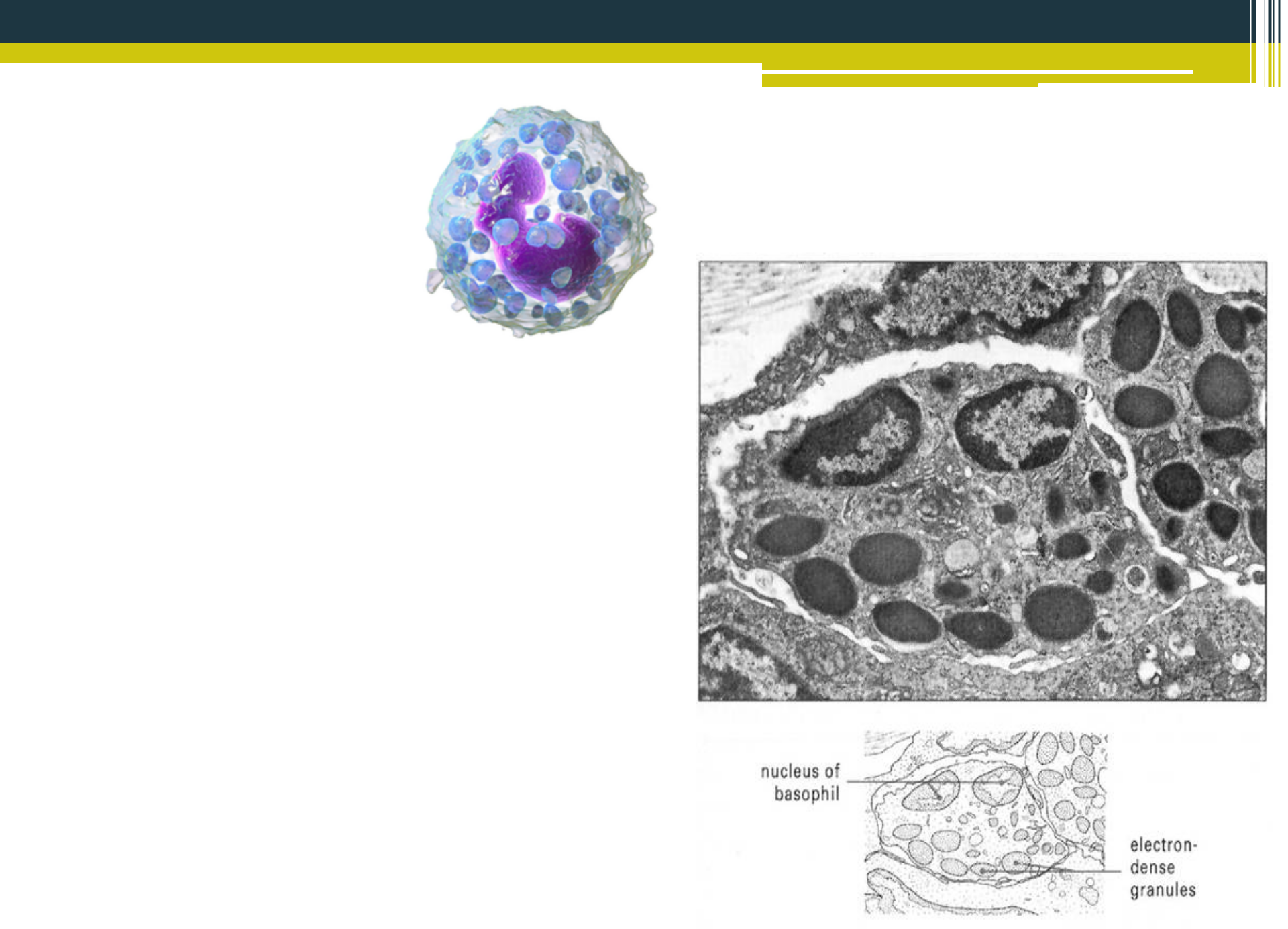{kind=link}
Mastocyty
Početne sú zastúpené v sliznici čreva, kože a pľúc. Mastocyty obsahujú receptory pre imunoglobulíny triedy G (Ig G) a zložky komplementu, čo im umožňuje pohlcovať opsonizované častice. Sú schopné priamej fagocytózy niektorých baktérií (hlavne enterobaktérií), pričom uvoľňujú mediátory zápalu (napr. histamín). K aktivácii mastocytov môže dôjsť aj väzbou Ig E molekúl, pričom dochádza k výraznej degranulácii biologicky aktívnych látok (histamín, serotonín, prostaglandín D2, leukotriény C4, D4), ktoré spôsobujú lokálnu bronchokonstrikciu, zvýšenú sekréciu hlienu, zvýšenú priepustnosť ciev. Žírne bunky môžu spôsobovať na slizničných bariérach výraznú indukciu zápalu, čo môže v niektorých prípadoch viesť k neprimeranej alergickej reakcii. Veľkosť mastocytov sa pohybuje v rozmedzí 10 - 20 pm, ich dĺžka života je 1 - 6 mesiacov.
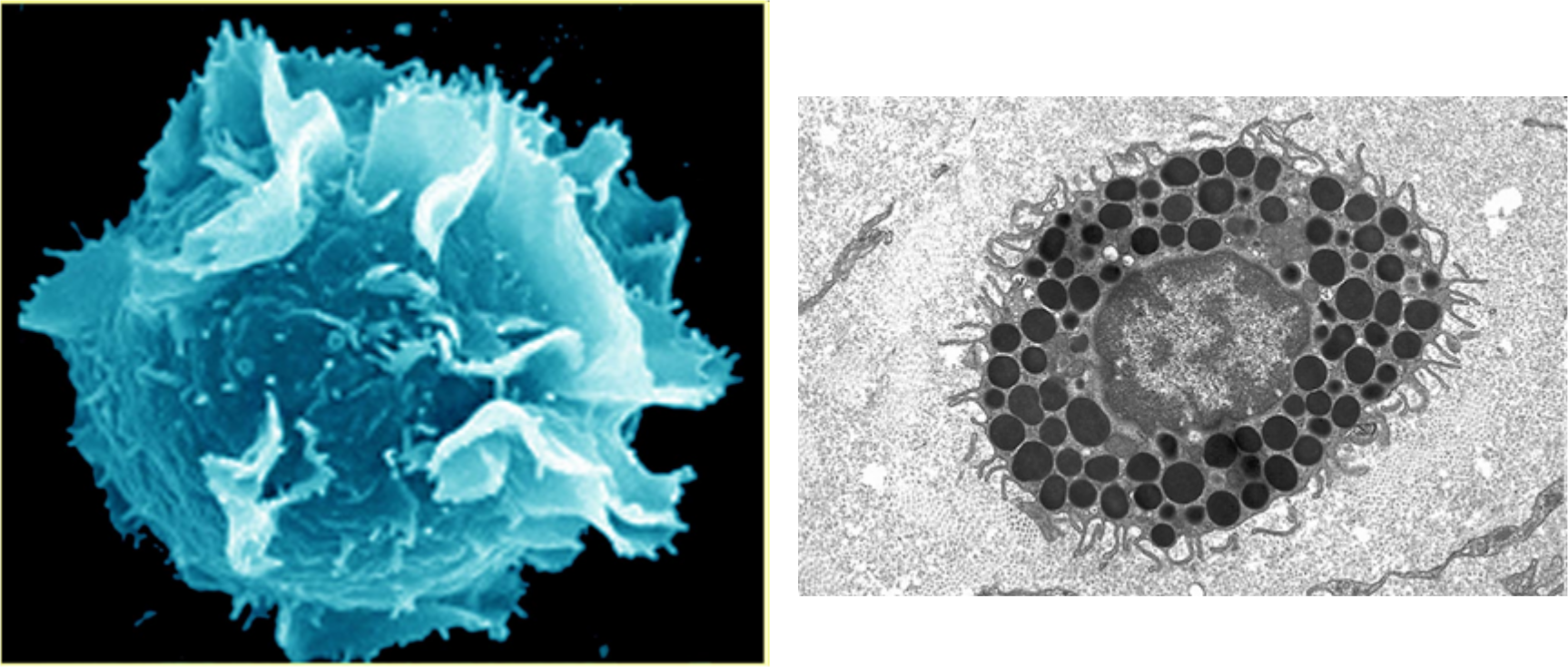{kind=link}
APC bunky
Osobitnú kategóriu buniek imunitného systému tvoria bunky prezentujúce antigén - APC bunky (antigen presenting cells), ktoré sú schopné upraviť cudzorodý antigén do takej formy, aby ho mohli rozpoznať T lymfocyty a zahájiť proti nemu špecifickú imunitnú odpoveď. Uvedená úprava sa týka najmä proteínových antigénov a jej podstatou je degradácia ich molekúl na imunogénne fragmenty, ktoré sa spájajú do komplexov s antigénmi hlavného histokompatibilného komplexu a až tento komplex sú schopné rozpoznať TH alebo TC lymfocyty. Na tento účel majú na svojom povrchu osobitné antigénové receptory, ktoré sa skrátene označujú TCR (T cell receptor). Medzi APC-bunky patria najmä makrofágy, monocyty, dendritické bunky, Langerhansove bunky kože, ale v prípade potreby aj endotelové bunky ciev, epitelové bunky čreva, niekedy aj B lymfocyty a ďalšie bunky.
Lymfoidná dráha
Z lymfoidnej dráhy vznikajú T lymfocyty, B lymfocyty a NK bunky. Lymfocyty vo všeobecnosti predstavujú 20 % zo všetkých leukocytov. Sú bunkami, ktoré podrobne rozpoznávajú a reagujú na cudzorodé antigény. V krvi cirkuluje asi len 5 % z celkového množstva lymfocytov, zvyšok sa nachádza najmä v slezine, lymfatických uzlinách, mandliach, Peyerových plakoch, týmuse a kostnej dreni. Lymfocyty tvoria rôznorodú populáciu buniek podľa pôvodu, štruktúry, povrchových antigénov a funkcie. Podľa veľkosti ich delíme na malé, stredné a veľké. V lymfatických tkanivách sú najviac zastúpené práve malé lymfocyty, tvoriace 80 - 95 % všetkých lymfocytov v týchto tkanivách. Sú to malé bunky guľatého tvaru s priemerom 6 - 8 um. Majú okrúhle, tmavo sa farbiace bazofilné jadro, obsahujúce najmä heterochromatín, ktoré je obklopené úzkym lemom cytoplazmy.
{kind=link}
Základné typy lymfocytov sú T- a B- lymfocyty a NK bunky. Zaraďujú sa k nim ešte aj K bunky a NKT bunky. Z lymfoidného prekurzora leukocytov vzniká 15 % B lymfocytov, 80 % T lymfocytov a 5 % tvoria spoločne NK bunky, K bunky a NKT bunky. Lymfocyty sa spoločne s monocytmi označujú ako mononukleárne bunky.
T lymfocyty
Podobne ako B lymfocyty, aj T lymfocyty vznikajú v kostnej dreni. Na rozdiel od nich, ale dozrievajú v týmuse (prebieha tu hlavná časť ich vývoja). T lymfocyty sú schopné regulovať aktivitu iných buniek a tiež zneškodňovať infikované bunky. Na rozdiel od B lymfocytov, nedokážu rozpoznať antigén priamo, t. j. antigén musí byť prezentovaný APC bunkami (napr. dendritovými bunkami, makrofágmi). T lymfocyty môžeme vo všeobecnosti rozdeliť na nasledovné populácie:
{kind=link}
- TH lymfocyty (pomocné) - s diferenciačným antigénom (povrchovým znakom) CD4+. Rovnakou mierou sa zúčastňujú na aktivácii procesov celulárnej a humorálnej imunity. Patria medzi kľúčové bunky aktivácie, diferenciácie a izotypovej regulácie B lymfocytov. Sú nevyhnutné pre aktiváciu B lymfocytov, ostatných T lymfocytov, makrofágov a NK buniek. TH lymfocyty takto tvoria základ celého imunitného systému. Pri ich poškodení alebo zničení (napr. vírusom HIV), dochádza k zrúteniu imunitného systému. V rámci tejto populácie, na základe odlišných znakov a funkčných vlastností, rozlišujeme nasledovné subpopulácie TH lymfocytov:
- TH0 - naivné, panenské bunky, ktoré neprišli do kontaktu s antigénom,
- TH1 - zápalové bunky, vyznačujúce sa cytotoxickou aktivitou,
- TH2 - bunky, ktoré „pomáhajú“ B lymfocytom rozpoznať antigén,
- TH3 - podieľajú sa na produkcii hojivých cytokínov.
- TR lymfocyty (regulačné) / TS lymfocyty (supresorové) - obsahujú diferenciačné antigény CD4+, CD25+. Zúčastňujú sa na procesoch riadenia imunitnej odpovede a vytváraní imunotolerancie. Ide o populáciu buniek, ktoré sa podieľajú na zabránení reakcií voči autoantigénom.
- TC lymfocyty (cytotoxické) disponujú povrchovými znakmi CD8+, CD45RO+, CD29+. Spoločne s TH lymfocytmi sú najpočetnejšou populáciou T lymfocytov. Podieľajú sa na kľúčových mechanizmoch celulárnej imunity ako efektorové bunky. Majú schopnosť cytotoxicky priamo usmrcovať vírusmi infikované bunky a nádorové bunky. V rámci tejto populácie buniek možno charakterizovať tzv. TM lymfocyty, ktoré sú dlhodobo schopné udržiavať antigénovú špecificitu T lymfocytov po prekonaní infekcie. Majú diferenciačný antigén CD450RO+.
- TI lymfocyty (iniciátorové) - iniciujú imunitnú odpoveď po kontakte s antigénom a vyvolávajú v regionálnej oblasti lymfatickej uzliny produkciu (a kumuláciu) lymfocytov. Prežívajú niekoľko mesiacov až rokov.
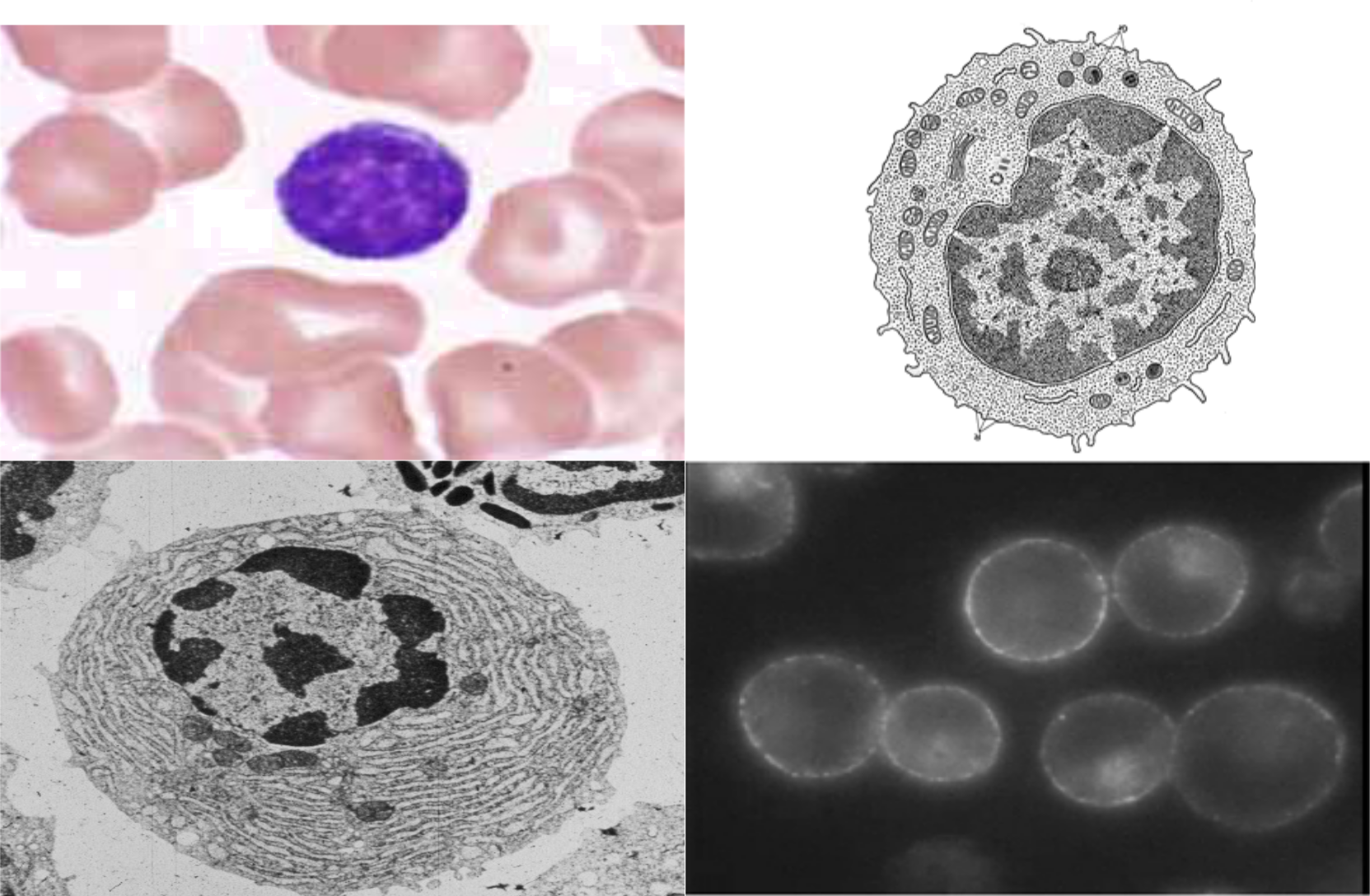
B lymfocyty
{kind=link}
Vývoj týchto buniek u cicavcov prebieha v kostnej dreni a dokončieva sa po stretnutí s antigénom v sekundárnych systémových lymfatických orgánoch (lymfatické uzliny, slezina), resp. v slizničnom imunitnom systéme (Peyerove plaky). U vtákov prebieha tvorba B lymfocytov vo Fabriciovom vačku.
Po stimulácii B lymfocytov antigénom sa ich syntéza zaktivizuje, pričom vznikajú imunoblasty, plazmoblasty a napokon plazmatické bunky (plazmocyty), ktoré sú hlavnými producentami protilátok (imunoglobulínov) v tele a konečným diferenciačným štádiom B lymfocytov. Plazmatické bunky obsahujú diferenciačný antigén CD 38, prežívajú len 3 - 6 dní. Morfologicky sú charakteristické excentricky uloženým jadrom a mohutnou retikulárnou formáciou, charakteristickou pre intenzívnu proteosyntézu. Jeden plazmocyt produkuje približne 300 molekúl imunoglobulínov za sekundu
Vývoj B lymfocytov môžeme rozdeliť na dve štádiá: na štádium nezávislé a na štádium závislé od prítomnosti antigénu v organizme. Štádium nezávislé od antigénu sa odohráva v kostnej dreni a predstavuje dozrievanie nezrelých B lymfocytov na zrelé, plne funkčné bunky. Odhaduje sa, že každý deň sa vyprodukuje približne 5 x 107 nových B lymfocytov, pričom do obehu sa dostáva len okolo 10 % z nich. Zvyšok je eliminovaný negatívnou a pozitívnou selekciou, pričom väčšinou ide o lymfocyty, ktoré sú citlivé na vlastné bunky.
Charakteristickým znakom pre tieto bunky je prítomnosť imunoglobulínov v cytosole a na cytoplazmatickej membráne. Tieto membránové imunoglobulíny sú tzv. antigén väzbovými receptormi. Väčšina B lymfocytov obsahuje na membránach Ig M a Ig D molekuly. V obehu je málo buniek nositeľmi Ig G, Ig A alebo Ig E molekúl, ale v črevnej sliznici sú Ig A pozitívne bunky v prevahe.
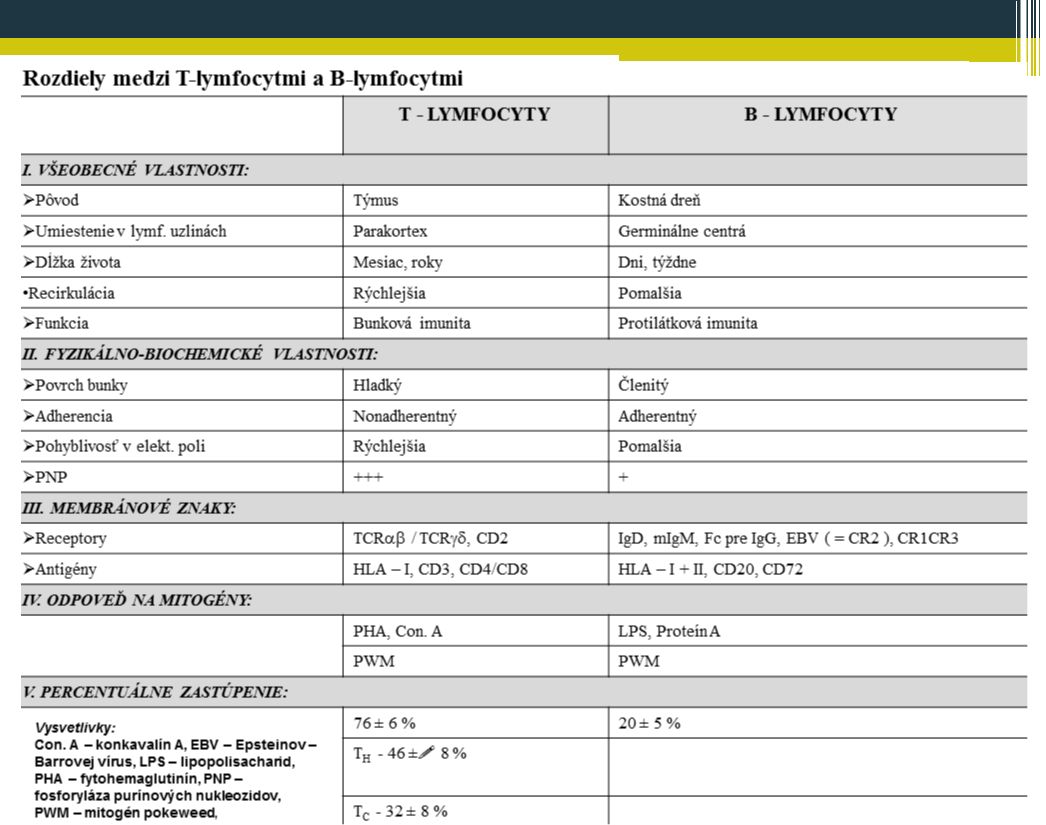
Rozdiely medzi T-lymfocytmi a B-lymfocytmi
{kind=link}
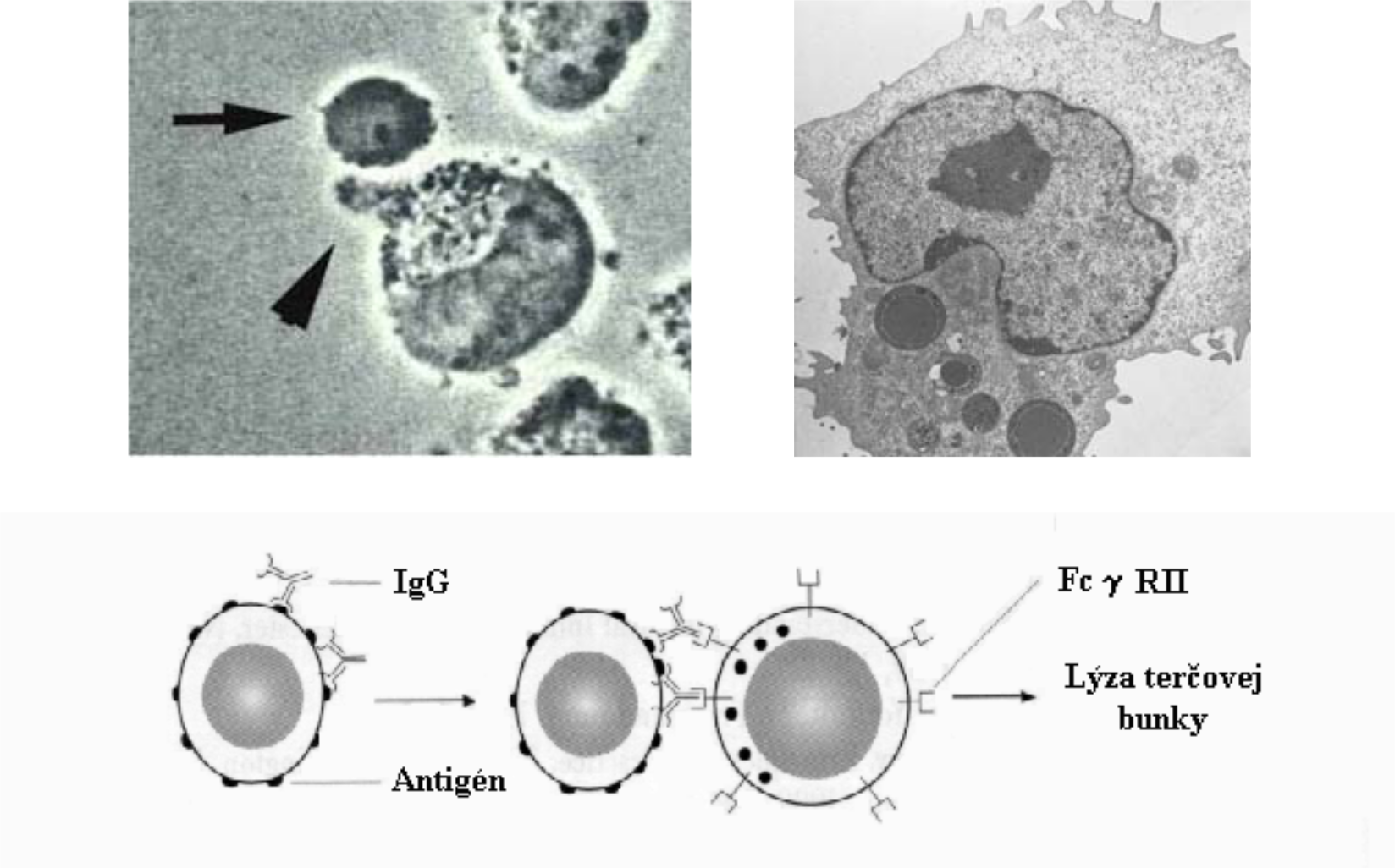
NK bunky
NK bunky (natural killer cells) sú veľkými granulárnymi lymfocytmi, ktoré nemajú antigénovo špecifické receptory ako B- a T- lymfocyty. Obsahujú diferenciačné antigény CD 16 a CD 56. Nešpecifickým spôsobom usmrcujú vlastné nádorovo transformované alebo vírusmi infikované bunky, ktoré majú na svojom povrchu málo MHC glykoproteínov triedy I (MHC I). Tieto bunky totiž potláčajú expresiu MHC I glykoproteínov, v dôsledku čoho sú chránené proti TC lymfocytom (TC lymfocyty totiž zneškodňujú takéto bunky). Z uvedeného dôvodu sa považujú za základné bunky prirodzenej obrany proti spontánne vznikajúcim nádorom a vírusom.
{kind=link}
Aj keď je ich funkcia podobná ako funkcia TC lymfocytov, na rozdiel od nich NK bunky nepotrebujú pre svoju činnosť stimuláciu a aktiváciu od konkrétneho antigénu a TH lymfocytov. NK bunky sa vyskytujú hlavne v krvi, slezine a produkujú cytokíny, podieľajúce sa na regulácii imunitných procesov. Aktiváciu NK buniek zabezpečujú adhezívne molekuly (napr. membránové lektíny C).
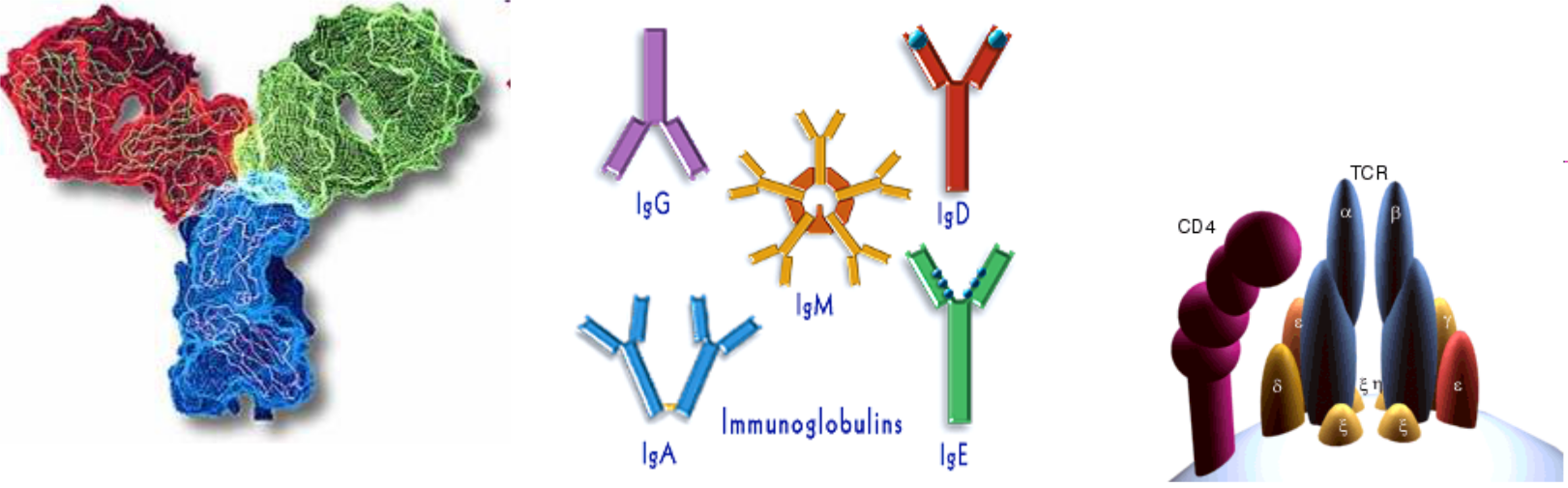
Molekuly imunitného systému
Molekuly imunitného systému majú dve základné funkcie: výkonnú a regulačnú. Výkonná funkcia sa prejavuje v bezprostrednej obrane proti patogénnym baktériám (niektoré toxické peptidy, voľné radikály odvodené od kyslíka sú schopné usmrcovať fagocytované baktérie). Regulačnú funkciu zabezpečujú regulačné molekuly (napr. imunohormóny, cytokíny), ktoré regulujú priebeh imunitných odpovedí, ale aj spoluprácu imunitného systému s ostatnými fyziologickými systémami organizmu.
{kind=link}
Z hľadiska svojich vlastností a funkcie môžeme molekuly pôsobiace v imunitnom systéme rozdeliť do niekoľkých skupín:
- protilátky (imunoglobulíny)
- cytokíny (napr. lymfokíny, interleukíny)
- imunohormóny (tymozíny)
- komplementový systém
- HLA molekuly (antigény) I. a II. triedy
- receptory - antigénovo špecifické receptory na povrchu T- a B- lymfocytov (TCR - T cell receptor, BCR - B cell receptor; FC receptory - viažu FC časti molekúl imunoglobulínov)