Imunobiológia
Obsah
Komplementový systém
Komplementový systém ( komplement ) bol objavený ešte koncom 19. storočia a predstavoval určitú súčasť čerstvého krvného séra, ktoré sa muselo pridať k suspenzii baktérií spoločne so špecifickou protilátkou, aby došlo k usmrteniu a lýze týchto baktérií. Pôvodný objaviteľ tohto javu Jules Bordet (Nobelová cena v roku 1919) nazval uvedenú súčasť séra ešte v roku 1896 alexín. Paul Ehrlich však neskôr zaviedol pre ňu termín komplement, čím vyjadril skutočnosť, že komplement dopĺňa (komplementuje) schopnosť protilátky lyzovať baktérie. Samotná protilátka túto schopnosť nemá. Neskôr sa ukázalo, že komplement nie je jedna látka, ale zmes viacerých látok.
{kind=link}
{kind=link}
V súčasnosti sa ako komplement označuje sústava viac než 30 rôznych glykoproteínov, prítomných v krvnej plazme, telových tekutinách a na povrchu niektorých buniek (kde utvárajú rôzne receptory). Tvorí asi 10 % proteínov plazmy. Väčšina zložiek komplementu sa syntetizuje v pečeni (až 90 %), niektoré v monocytoch, makrofágoch, neutrofiloch, fibroblastoch a iných bunkách. Táto lokálna syntéza komplementu umožňuje rýchly rozvoj zápalového procesu v extravaskulárnych priestoroch. Zložky komplementového systému sa označujú písmenom C, ku ktorému je priradené číslo podľa poradia, v akom sa jednotlivé zložky postupne objavovali. Niektoré zložky nie sú očíslované, ale označujú sa ako faktory, napr. faktor B, faktor D, faktor P (properdín). Patria k alternatívnej ceste aktivácie komplementu a regulačným proteínom. Iné zložky komplementu majú svoje označenie podľa funkcie, ktorú vykonávajú, napr. MBL (mannose binding lectin - proteín viažúci sa na manózu).
Všetky zložky komplementu (okrem faktora D) sa syntetizujú v neaktívnom stave. Aktivácia komplementu je kaskádovitý proces, keď mechanizmom limitovanej proteolýzy sa z pôvodného neaktívneho prekurzora odštiepením určitých úsekov proteínu a tým odkrytím reaktívneho miesta stane enzýmovo aktívny fragment, ktorý môže štiepiť ďalšiu zložku a tým ju aktivovať. Takto sa aktivita komplementu amplifikuje. Vzniká tak určitý „biologický vodopád“ s mnohostrannými biologickými účinkami, dokonca s možným negatívnym dopadom na organizmus. Práve preto sa zložky komplementu syntetizujú vo forme neaktívnych proenzýmov a ich aktivácia je starostlivo regulovaná.
{kind=link}
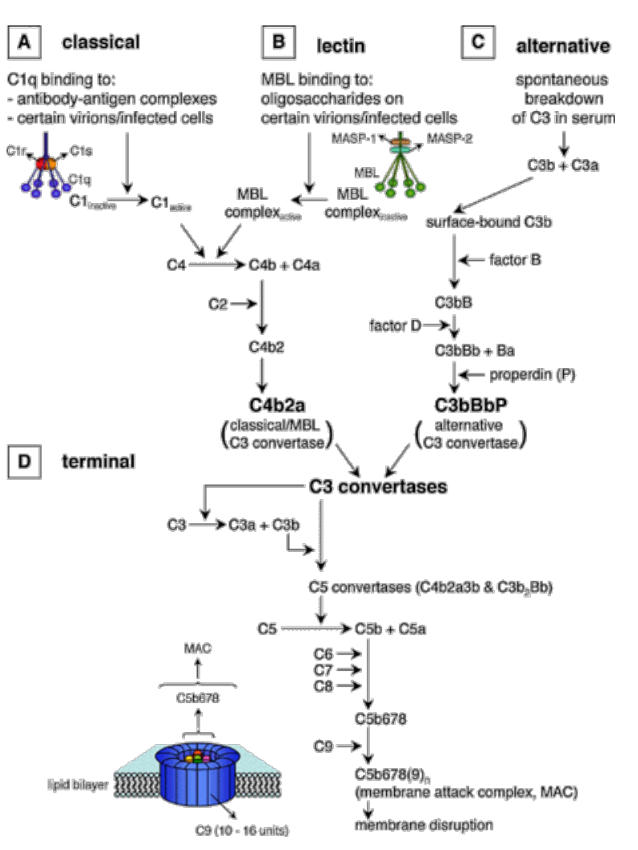
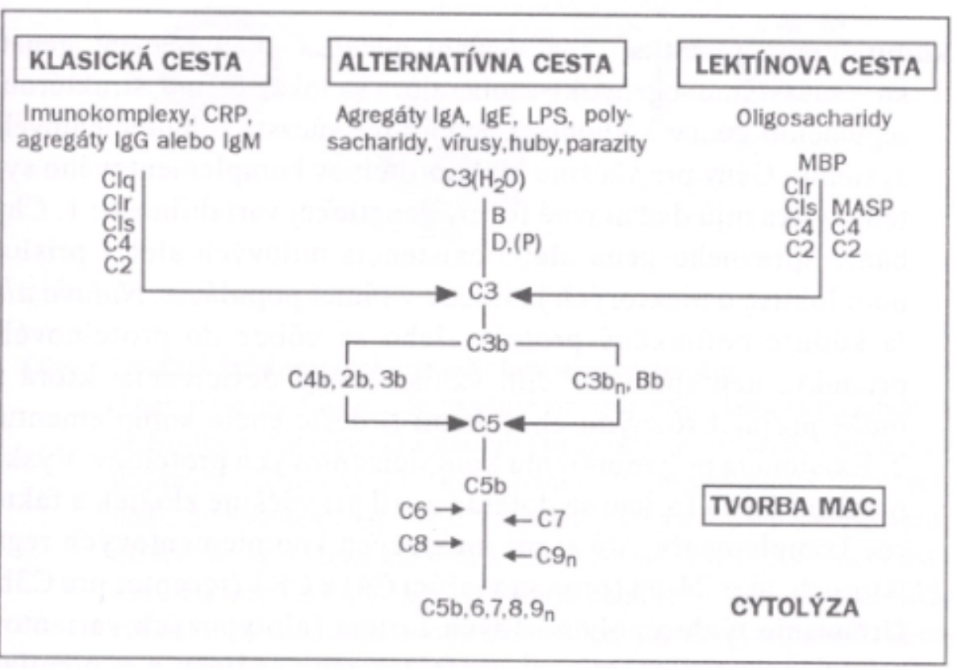
Zložky komplementu sa môžu aktivovať tromi rôznymi cestami:
- imunokomplexmi alebo C-reaktívnym proteínom (klasická cesta aktivácie);
- niektorými sacharidmi a lektínmi (lektínová cesta aktivácie);
- inými látkami väčšinou mikróbneho pôvodu (alternatívna cesta aktivácie).
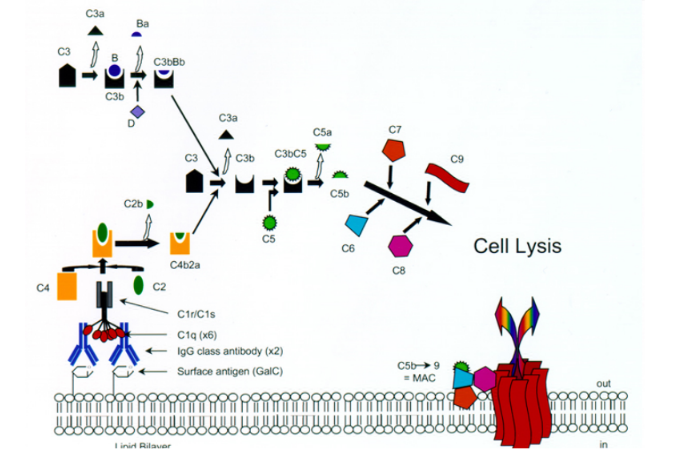
Hlavnú zložku komplementu predstavuje 9 sérových proteínov (C1-C9). Ústrednou zložkou je proteín C3, ktorého fragment C3b sa kovalentne viaže na mikrobiálny povrch (vzniká pri každej ceste aktivácie komplementu). Pri rôznych podnetoch dochádza ku kaskádovitej aktivácii jednotlivých zložiek, medziprodukty tejto reakcie majú výrazné biologické funkcie (opsonizácia, chemotaxia). Terminálny (konečný) produkt komplementovej kaskády predstavuje komplex proteínov C5b678(9)n (označovaný ako MAC - membrane attack complex, čiže komplex atakujúci membrány), ktorý perforuje membrány niektorých mikroorganizmov, spôsobuje ich lýzu a tým ich zneškodňuje.
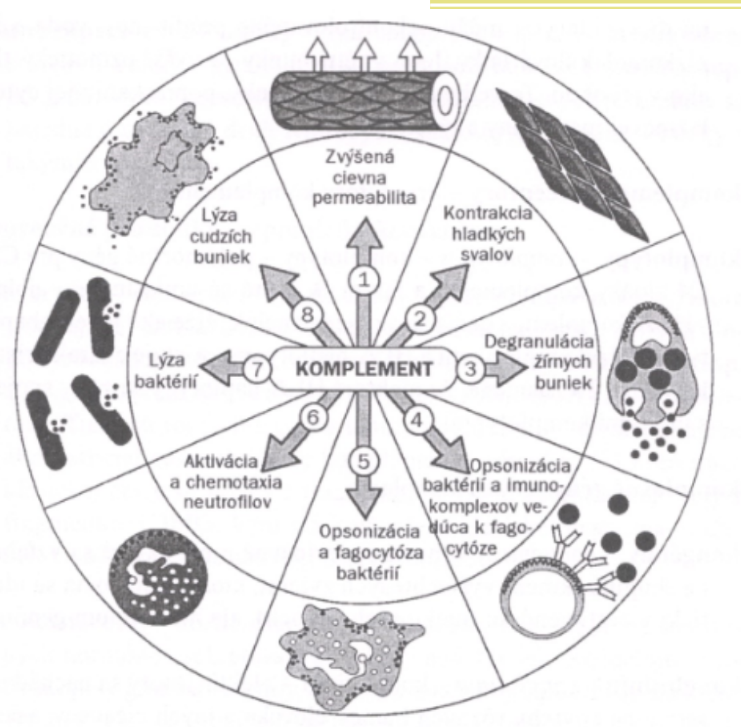
Hlavné funkcie komplementového systému sú teda:
- opsonizácia (C3b);
- chemotaxia (C3a, C5a);
- osmotická lýza (komplex C5b - C9).
Niektoré zložky komplementového systému sú však významné i pri iných dejoch, napr. imunokomplexy obsahujúce fragmenty C3b regulujú transport antigénov do sleziny a lymfatických uzlín a niektoré komplementové receptory slúžia aj ako adhezívne molekuly.
Aktivácia komplementu
Komplementový systém sa aktivuje 3 spôsobmi a to: alternatívnou, klasickou a lektínovou cestou.
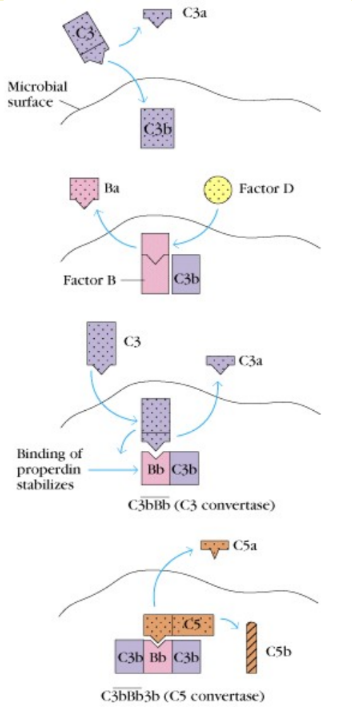
Alternatívna cesta aktivácie komplementu je v pravom zmysle slova nešpecifickou imunitnou reakciou (aktivuje sa okamžite po vstupe cudzorodého infekčného zárodku alebo aj iných látok do organizmu; spúšťajú ju niektoré polysacharidy, lipopolysacharidy, veľké vírusy, niektoré mikroorganizmy a bunky niektorých nádorov). Kľúčová zložka komplementu - proteín C3 sa s nízkou frekvenciou samovoľne štiepi na väčší fragment C3b a menší C3a. Vo vzniknutom C3b sa odhalí veľmi reaktívna cyklická tioesterová skupina, ktorá bola v intaktnej molekule skrytá. Táto skupina rýchlo reaguje s hydroxy- a aminoskupinami v blízkom okolí. Pokiaľ sú to skupiny na povrchu nejakej častice (bunky vlastné organizmu, mikroorganizmus), naviaže sa touto reakciou C3b kovalentne na takýto povrch. Molekula C3b naviazaná na povrchu častice zaháji kaskádu ďalších reakcií: najprv sa k nej pripojí ďalší sérový proteín (faktor B) a ten je potom štiepený sérovou proteázou (faktor D) na fragmenty Ba a Bb. Vzniknutý komplex C3bBb je stabilizovaný faktorom P (properdín) a pôsobí ako tzv. alternatívna C3 konvertáza (enzým štiepiaci veľmi účinne C3 na C3a a C3b. Vzniknuté fragmenty C3b sa kovalentne zachytávajú na povrchu okolo enzýmovo aktívneho komplexu, ktorý tento dej vyvolal a slúžia ako významné opsoníny. Okrem toho z niektorých vznikajú ďalšie molekuly C3 konvertázy a tak dochádza k mnohonásobnému zosilňovaniu pôvodného podnetu. Fragmenty C3a pôsobia chemotaxicky na fagocyty. Z niektorých molekúl C3 konvertázy vznikajú zložité komplexy (C3bBbC3b), ktoré majú odlišnú proteolytickú aktivitu (štiepia proteín C5 na fragmenty C5b a C5a). C3bBbC3b je teda alternatívnou C5 konvertázou. Vznikom C5b začína terminálna (lytická) fáza komplementovej kaskády. Fragmenty C5b vytvoria komplex s ďalšími zložkami komplementu (C6, C7, C8). Uvedený komplex (C5b678) sa ponorí do povrchovej lipidovej membrány atakovanej bunky a pripojí sa k nemu do kruhu 10-18 molekúl C9. Týmto spôsobom vzniknú v membráne póry, ktorými môžu unikať cytoplazmatické komponenty, poruší sa osmotická rovnováha (vo vnútri buniek je vyšší osmotický tlak ako v mimobunkovom prostredí, čo má za následok zväčšovanie objemu bunky až napokon prasknutie jej cytoplazmatickej membrány) a bunka podlieha lýze (uvoľnenie vnútorného obsahu).
{kind=link}
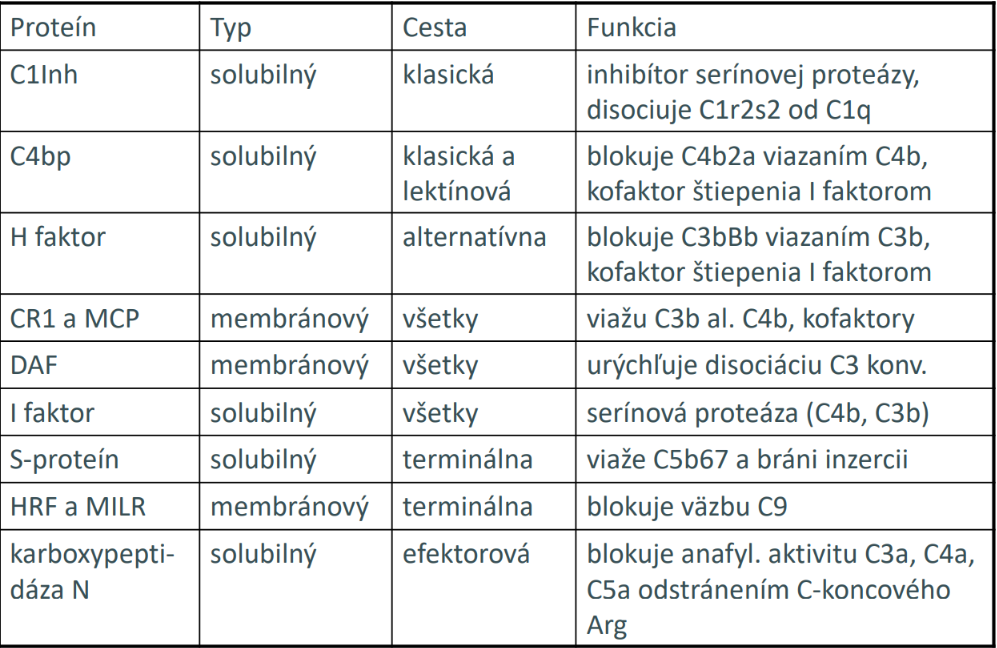
Niektoré mikroorganizmy sú však voči lytickému pôsobeniu komplementu odolné (ochrana bunkovou membránou). Uvedené procesy sú iniciované spontánnym, nešpecifickým spôsobom (štiepenie C3) a v princípe môže dôjsť k ich rozvoju ako na povrchu cudzorodých častíc, tak aj na povrchu vlastných buniek (škodlivé). V prípade týchto buniek sa však na povrchu vyskytuje niekoľko ochranných regulačných proteínov, brániacich rozvoju kaskády samovoľných reakcií.
Klasická cesta aktivácie komplementu sa uskutočňuje na povrchoch antigénov, na ktoré sa naviazali protilátky, čiže spúšťajú ju najčastejšie imunokomplexy (komplexy antigénu a špecifickej protilátky triedy Ig G alebo Ig M). Väzba protilátky na antigénový povrch spôsobí zmenu v konformácii jej molekuly tak, že sa odhalí väzbové miesto pre proteín C1. Ten po naviazaní sa na protilátku taktiež zmení svoj tvar a nadobudne proteolytickú aktivitu - začne štiepiť proteíny C4 a C2. Ich fragmenty C4b a C2a sa naviažu na povrch napadnutého mikroorganizmu a vytvoria klasickú C3 konvertázu (C4bC2a). Uvedený enzým štiepi veľké množstvo C3 na C3a a C3b, ktoré pôsobia rovnakým spôsobom ako v prípade alternatívnej cesty aktivácie komplementu. Neskôr sa vytvára ďalší enzým - klasická C5 konvertáza (C4bC2aC3b), ktorý štiepi C5 na C5b a C5a.
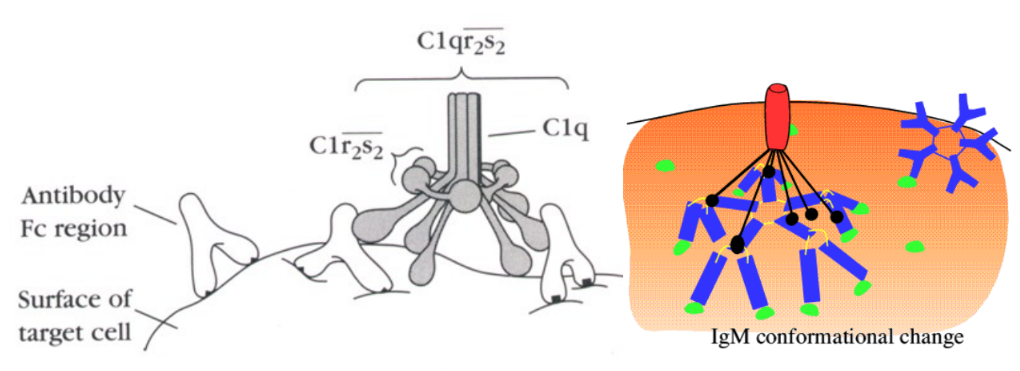{kind=link}
Klasickú cestu môžu okrem protilátok zahájiť taktiež tzv. pentraxíny (C-reaktívny proteín a sérový amyloid P). Tieto sérové proteíny (produkované hepatocytmi) sa viažu na niektoré mikroorganizmy a následne sú schopné viazať a aktivovať proteín C1.
Nevýhodou klasickej cesty aktivácie komplementu je časový posun jej naštartovania, a to 4 - 5 dní (potrebné na vytvorenie špecifických protilátok). Organizmus si však v určitých nevyhnutných situáciách vie pomôcť a urýchliť nástup klasickej cesty aktivácie, napr. počas akútnej zápalovej fázy (v priebehu 1 - 2 dní) vzniká C-reaktívny proteín, ktorý môže zahájiť aj klasickú cestu aktivácie. Takýmto spôsobom uvedený proteín do určitej miery vykrýva obdobie nevyhnutné na tvorbu špecifických protilátok.
Lektínová cesta aktivácie komplementu je podobná klasickej ceste, ale namiesto protilátok je iniciovaná sérovým lektínom (MBL - mannose binding lectin, čiže proteín syntetizovaný hepatocytmi a viažúci sa na manózu). MBL sa viaže na manózu (sacharid nachádzajúci sa na povrchu buniek v glykokalyxe) niektorých mikroorganizmov priamo (nezávisle na protilátkach) a jeho štruktúra a funkcia je podobná C1. Po väzbe na mikrobiálny povrch taktiež štiepi C4 a C2.
V porovnaní s klasickou cestou sa môže aktivovať okamžite bez predchádzajúcej potreby indukcie tvorby špecifických protilátok.
Funkcie komplementu
{kind=link}
Komplementový systém je základným humorálnym mechanizmom prirodzenej imunity a niektorými svojimi aktivitami je prepojený s ostatnými tromi multienzýmovými systémami krvnej plazmy (hemokoagulačný, fibrinolytický a kinínový systém). Primárne ovplyvňuje aktivitu mnohých buniek a tkanív, ako aj priebeh viacerých fyziologických a patofyziologických procesov v organizme človeka a ostatných cicavcov.
Aktivácia komplementovej kaskády s tvorbou komplexu atakujúceho membrány má za následok rôzne cytotoxické a cytolytické reakcie. Výsledok cytotoxických reakcií môže byť pre organizmus prospešný (likvidácia infekčných agens alebo poškodených, či odcudzených buniek) alebo škodlivý (poškodenie vlastných buniek v imunopatologických reakciách). Pri aktivácii komplementu okrem cytotoxického MAC vznikajú aj rôzne fragmenty s ďalšími významnými biologickými aktivitami:
- anafylatoxíny - C5a, C3a, C5adeArg (molekula C5a bez N-koncového arginínu), C4a, ktoré vyvolávajú zvýšenú priepustnosť ciev, kontrakcie hladkých svalov a degranuláciu žírnych buniek a bazofilov. Sú to polypeptidové reťazce zložené zo 75-tich aminokyselinových jednotiek a spĺňajú všetky kritériá pre lokálne hormóny;
- chemotaktické faktory - C5a, C5adeArg - majú aktivačný a chemotaktický účinok najmä na neutrofily;
- opsoníny -obaľujú baktérie, imunokomplexy a iné častice, čím uľahčujú ich fagocytózu profesionálnymi fagocytmi, ktoré majú na svojom povrchu receptory pre tieto fragmenty (CR1, CR3);
- stimulátory respiračného vzplanutia profesionálnych fagocytov - C3b, C5a, C5adeArg. Pri respiračnom vzplanutí vznikajú rôzne cytotoxické formy kyslíka, ktoré sa zásadným spôsobom zúčastňujú na usmrcovaní fagocytovaných mikroorganizmov, ale aj na poškodzovaní vlastných buniek a tkanív pri poškodzujúcich zápalových reakciách.
Fragmenty C3b sa spolu s komplementovými receptormi CR1 a CR3 zúčastňujú aj na solubilizácii cirkulujúcich imunokomplexov. Cirkulujúce imunokomplexy sa pritom obalia molekulami C3b a tento komplex sa potom naviaže na uvedené receptory (najmä na erytrocytoch a neutrofiloch), čím sa odstránia z cirkulácie. To zabráni, aby sa usadili v tkanivách a vyvolali imunokomplexové ochorenia.
Z funkčného hľadiska môžeme komplementové receptory rozdeliť na dva typy: adherenčné a ostatné. Adherenčné receptory sprostredkúvajú adhéziu buniek a iných častíc, obalených C3b alebo C4b fragmentami a označujú sa ako CR1 až CR5. Adhezívna reakcia sprostredkovaná CR na fagocytoch má za následok stimuláciu fagocytózy, aktiváciu kyslíkového metabolizmu a sekrečných funkcií fagocytov. Druhý typ komplementových receptorov reaguje s malými fragmentami niektorých zložiek (C5a, C3a, C4a) alebo faktorov (Ba, Bb), prípadne niektorých regulačných proteínov. Stimulácia týchto receptorov má za následok rôzne biologické účinky (chemotaxiu, sekréciu vazoaktívnych amínov, mediátorov zápalovej a anafylaktickej reakcie a pod.).
{kind=link}