Fyziológia bunky
Obsah
Centrozóm
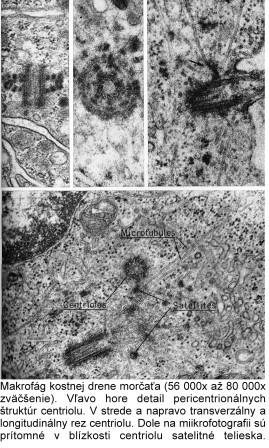
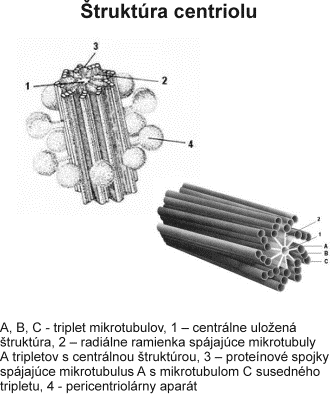
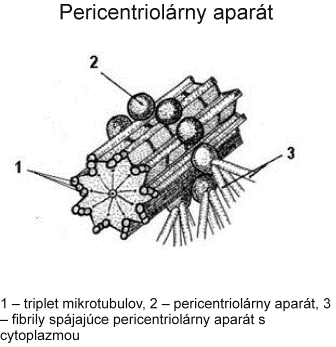
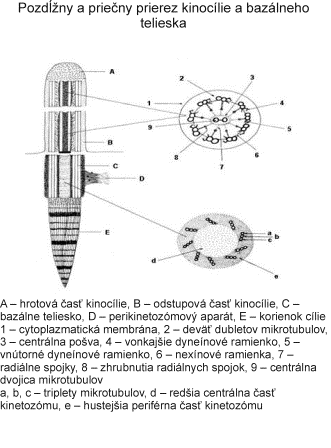
Centrozóm tvorí centriol, centrosféra a astrosféra. Je to vlastne dvojica centriolov usporiadaná kolmo k sebe (diplozóm) a okolitá cytoplazma. Centriol je teliesko tvaru valca o veľkosti 0,4 – 0,5 µm a šírke asi 0,1 – 0,2 µm. Vyskytuje sa takmer vo všetkých typoch buniek a je lokalizovaný najčastejšie v blízkosti jadra alebo Golgiho aparátu. Vo vonkajšej časti prebieha pozdĺžne 9 navzájom spojených trojíc vysoko organizovaných mikrotubulov, usporiadaných do kruhu, označovaných triplety. Každá trojica má vnútorný tubulus A úplný, tubuly B a C nasadajú na tubulus A, ale nie sú kompletné.
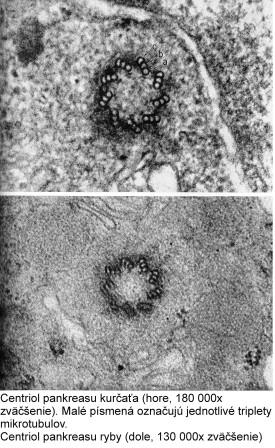{kind=link}
{kind=link}
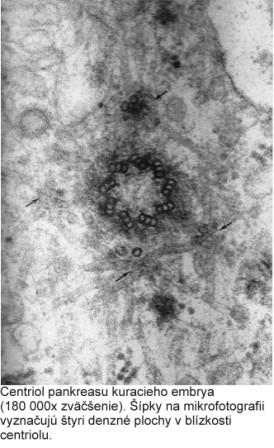{kind=link}
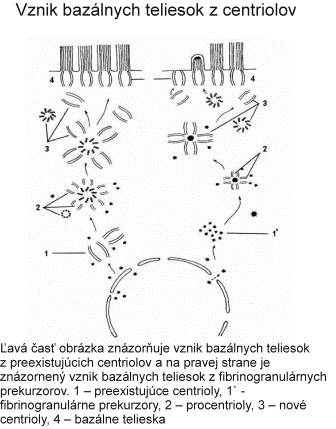
Centrozóm má veľký význam aj pri tvorbe bazálnych teliesok (kinetozómov) riasiniek a bičíkov. Pri vzniku nových riasiniek prebieha autoreprodukcia centriolov, ktoré sa presúvajú k povrchu bunky a stávajú sa kinetozómami vyrastajúcich riasiniek. Po ukončenií diferenciácie riasiniek plnia opäť úlohu ich kinetického centra. Podobný proces prebieha aj pri vzniku bičíkov. Obsah DNA v centriole podmieňuje schopnosť množenia (reduplikácie, autoreplikácie) centriolov. Proces rozmnožovania však neprebieha ako rozdvojenie na dva dcérske centrioly, ale ako mechanizmus zahŕňajúci utvorenie tzv. procentriolu v blízkosti materského centriolu. Procentriol uložený kolmo k pôvodnému centriolu rastie a vzniká zrelý centriol.
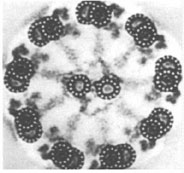{kind=link}
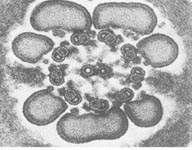{kind=link}
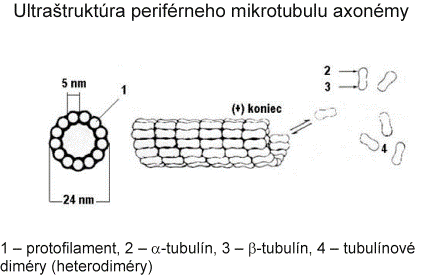
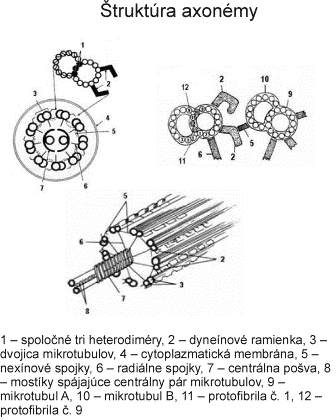
Bičíky (flagelá) a riasinky (cílie) majú veľmi podobnú štruktúru ako centriol. Rozdiel spočíva v tom, že v bazálnom teliesku, ktoré vzniklo z centriolu sa nachádza 9 periférnych subjednotiek (triplety), ale v bičíku a riasinkách sa nachádzajú okrem 9 periférnych subjednotiek (dublety + dyneínové ramienka) aj 2 centrálne mikrotubuly, ktoré sa utvárajú až v úrovni povrchu bunky (sú viditeľné iba nad povrchom bunky). Bazálne telieska slúžia ako matrice kontrolujúce usporiadanie axonemálnych podjednotiek.
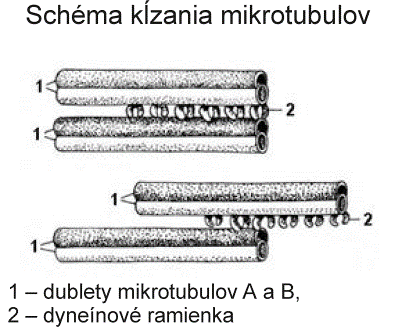
Keďže základný mechanizmus pohyblivosti je rovnaký na všetkých pohyblivých bunkových výbežkoch, ktoré majú usporiadanú axonému podľa vzorca 9 + 2, závery o mechanizme pohyblivosti platia pre cílie i bičíky spermií. Cílie kmitajú v skupinách v takom časovom slede, ktorý umožňuje posúvanie membránovej vrstvy hlienu jedným smerom po povrchu buniek. Pohyb bičíkov i riasiniek zabezpečuje interakcia dyneínu a tubulínu periférnych mikrotubulov. Pri kontrakcii dyneínové ramienka obopnú B mikrotubuly susedných dubletov, posúvajú sa pozdĺž povrchu B mikrotubulu susedného dubletu, priemer bičíka (riasinky) sa dočasne zníži a táto kontrakčná vlna sa posúva po celej dĺžke bičíka alebo riasinky. Kĺzavý pohyb medzi priľahlými mikrotubulmi obmedzuje prítomnosť nexínu a radiálnych lúčov, takže sily vyvinuté týmto procesom riasinky a bičíky ohýbajú, čím sa vyvolá ich pohyb. Po kmite sa uvoľnia, ohnú sa pod hlienovou vrstvou, aby ju neporušili alebo neposunuli späť a vracajú sa do východiskovej polohy. Frekvencia pohybu cílie je 2,5 kmitu za sekundu. Zdrojom energie je adenozíntrifosfát.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}